
- Отдел моховидные, их строение и происхождение
- Особенности строения и развития моховидных
- Жизненный цикл маршанции
- Жизненный цикл зеленого мха кукушкин лен
- Вегетативное размножение мхов
- Важнейшие особенности моховидных
- Симбиоз мхов и насекомых
- Цикл развития папоротникообразных растений на примере папоротника щитника мужского
Отдел моховидные, их строение и происхождение
 |




Классификация растений, их характеристика и жизненные циклы
Мхи или моховидные – отдел высших растений, объеденных в классы: печёночные мхи (6 000–8 000 видов), антоцеротовые мхи (100–200 видов) и листостебельные мхи (10 000 видов). Мхи это, как правило, мелкие растения, длина которых редко превышает 50 мм. Отличаются от других высших растений тем, что в их жизненном цикле взрослые растения представлены разнополыми гаметофитами, а спорофит развивается на женском гаметофите. Размножаются бесполым путём – спорами и вегетативно частями таллома.
В цикле развития взрослые (многолетние) растения маршанции полиморфной представлены разнополыми (двудомными) мужскими и женскими гаметофитами. Вегетативное тело растения – пластинчатый таллом, имеет вид мясистого лопастного, дихотомически разветвленного слоевища длиной до 10-12 см и шириной до 3 см, образованного множеством клеточных слоев (около 30). Специализированных проводящих тканей таллом не имеет. На нижнем эпидермисе образуются два типа выростов:
– ризоиды – бесцветные простые и язычковые одноклеточные выросты, которые выполняют функцию закрепления растения в субстрате. Воду маршанция способна поглощать всем телом,
– амфигастрии – окрашенные (темно-фиолетовые) однослойные, многоклеточные чешуи (редуцированные «листья» – филоиды), которые способны долго удерживать воду во время засухи.
Различить мужской и женский гаметофиты возможно во время развития на поверхности талломов специальных органов (подставок для гаметангиев) – мужских антеридиефоров и женских архегониефоров.
Антеридиефор представляет собой ножку с расположенным сверху восьмиугольным диском (шапкой). С верхней стороны шапки в антеридиальных камерах находятся мужские гаметангии – антеридии. В антеридиях образуются мужские двужгутиковые гаметы – сперматозоиды.
Архегониефоры развиваются на женских гаметофитах в виде девятилучевого щитка (звездочки) на ножке. Во время развития архегониефора, архегонии (женские гаметангии) закладываются с морфологически верхней стороны подставки. В силу неравномерного роста обеих сторон ложа подставки (щитка) они смещаются между лучами щитка на его нижнюю сторону, где располагаются группами. Вокруг каждой группы архегониев образуется специальная защитная обертка. Архегонии представляют собой кувшиноподобные камеры, направленные горловиной вниз, в брюшке которых образуется по одной яйцеклетке.
Половой процесс в цикле развития маршанции происходит посредством воды преимущественно во время дождя. Капли дождя выбивают из антеридиев зрелые сперматозоиды и вместе с ними отражаются на женские шапки архегониефоров. Сбегая между лучами женских подставок, капли воды вместе со сперматозоидами зависают в месте расположения архегониев, покрывая их. По воде мужские гаметы проникают через шейку архегония в брюшко – калиптру, где оплодотворяют яйцеклетку, образуя зиготу (первую диплоидную клетку спорофита). После оплодотворения вокруг каждого архегония из его ножки начинает образовываться индивидульная чашевидная защитная обертка – 4-5 лопастной псевдоперианций. В это время зигота делится митозом, формируя зародыш спорофита, который прикрепляется к материнскому телу (щитку архегониефора) присоской – гаусторией, и получает оттуда питание. С увеличением размеров зародыша калиптра (брюшко архегония) растягивается (увеличивается в размерах).
Взрослый спорофит, развивающийся из зародыша, включает в себя: стопу (присоску) – закрепляющую часть, ножку и спорангий (коробочку спорогона). В процессе своего роста спорофит разрывает калиптру и выносит спорангий наружу. В спорангии формируются диплоидные клетки спорогенной ткани, которые получают питание через удлиненные клетки со спирально утолщенными клеточными стенками – элатеры. В результате разделения спорогенных клеток мейозом образуются гаплоидные гетероталличные споры. При созревании спор элатеры высыхают и скручиваются спирально, разрыхляя массу спор в спорангии. Однослойная коробочка спорангия при созревании спор легко раскрывается восемью загнутыми назад зубцами, а пружинки элатер способствуют равномерному рассеиванию спор. Гетероталличные (разнополые) споры маршанции в благоприятных условиях прорастают в протонемы (передростки) – небольшие беспигментные нитчатые выросты, с верхушечной клетки которых образуются талломы соответственно мужского и женского гаметофита нового поколения.
Вегетативное размножение маршанции происходит с помощью выводковых почек, которые формируются в специальных выростах – выводковых корзинках, на верхней стороне как женских, так и мужских талломов. В результате попадания воды на таллом выводковые почки вымываются (распрыскиваются) и прорастают на почве в новые талломы соответствующего пола. Маршанция распространена в увлажненных местах на берегах водоемов, под покровом леса.
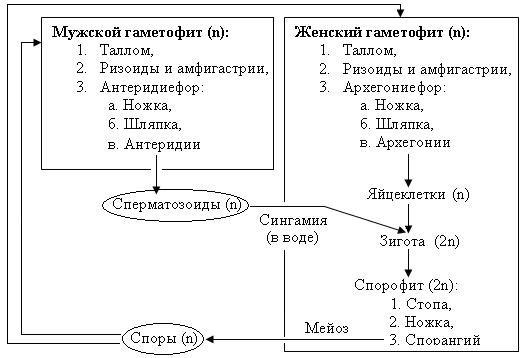
Рис. 30. Схема жизненного цикла мха маршанция
Спорофитплауна булавовидного – многолетнее вечнозеленое растение, преобладает в цикле развития. В своем составе имеетстелющийся дихотомически разветвленный стебель, покрытый спирально расположенными маленькими ланцетно-линейными листочками (микрофиллами) и закрепленный в почве дихотомически разветвленнымикорнями. Стеблевые побеги заканчиваются верхушечнымипочкамиили спороносными колосками (стробилами). Спороносные колоски на длинных ножках состоят изоси, на которой спирально расположены спороносные листья (спорофиллы) со спорангиями с верхней (дорсальной) стороны. После редукционного (мейотического) деления материнских диплоидных клеток образуются гаплоидныеспоры. Споры покрыты двойной оболочкой (эндо- и экзоспорием) и содержат до 50% масла. После созревания споры рассеиваются и в благоприятных условиях прорастают. Развитие гаметофита (заростка) из спор происходит медленно в течение 12-20 лет. Он образуетризоиды, которыми закрепляется в почве и поглощает воду, а также вступает всимбиозс мицелием гриба, который размещается в его коровой части. Заросток развивается в почве без доступа света, поэтому не имеет хлоропластов, однако если он выходит на поверхность, они образуются.Гаметофитдвуполый и напоминает луковицу, позже разрастается до 2-3 см в диаметре, и имеет блюдцеобразную форму.Антеридиииархегонииразмещены рядом на верхней стороне и погружены в паренхимную ткань. Антеридии овальной формы, архегонии – колбообразные. В брюшной части архегониев содержитсяяйцеклеткаи брюшная канальцевая клетка, в шейке – шейные канальцевые клетки. Для перемещениясперматозоидовс антеридиев к яйцеклеткам архегониев необходима вода. Заплывая в шейку архегония, сперматозоид движется в брюшко архегония, где сливается с яйцеклеткой, образуязиготу– первую диплоидную клетку спорофита.Зародыш– молодой спорофит, медленно развивается на гаметофите, получая из него необходимые питательные вещества. Когда корни зародыша прорастают в почву, он отсоединяется от гаметофита и разрастается во взрослое растение. При постепенном созревании архегониев на гаметофите в них могут образовываться и развиваться несколько разновозрастных спорофитов одновременно.
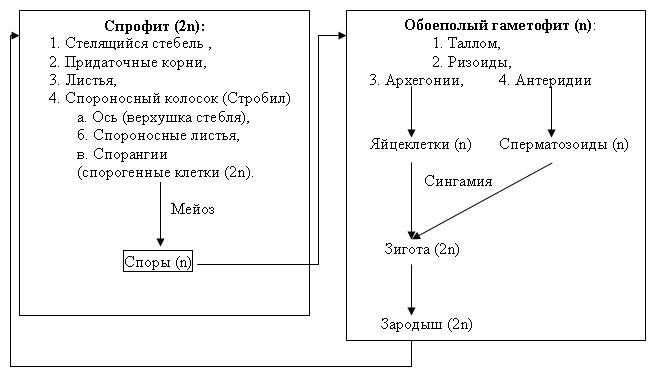
Рис. 36. Схема жизненного цикла плауна булавовидного
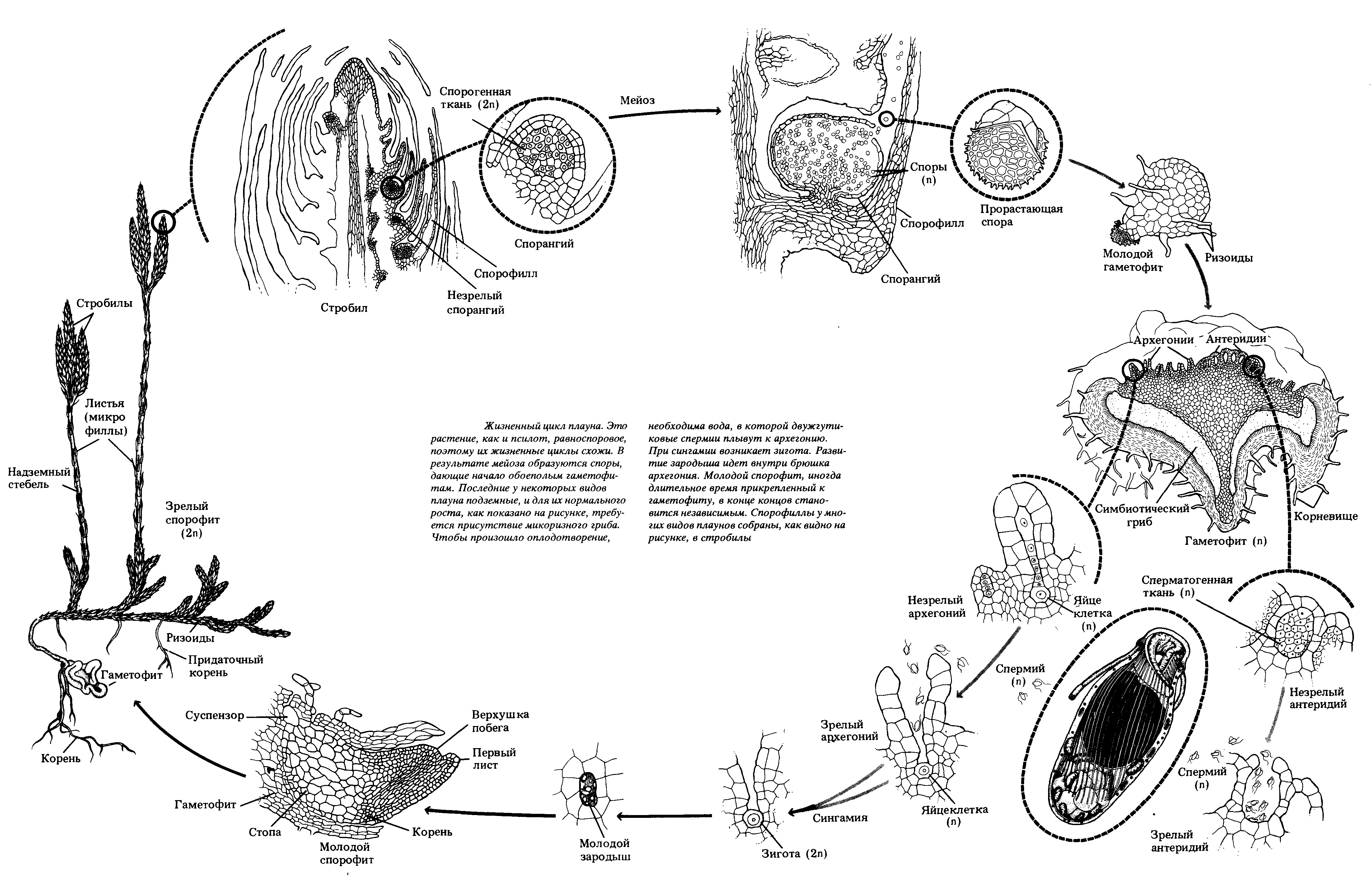
Рис. 37. Жизненный цикл плауна булавовидного
Папоротникообразные– отдел высших споровых растений, объединяющий около 11 000 видов. Папоротникообразные растения характеризуются корнепобеговым строением тела спорофитов и талломным строением тела гаметофитов. Размножаются бесполым путём – спорами и вегетативно – побегами (корневищами).
Цикл развития папоротникообразных растений на примере папоротника щитника мужского
В цикле развития папоротника преобладает спорофит. Взрослый многолетний спорофит (растение с диплоидным набором хромосом) представляет собойкорневище, закрепленное в почвепридаточными корнями. Из почек корневища весной на поверхность почвы выходят спороносные листья –вайи, с нижней стороны которых располагаютсяспорангии, собранные группами всорусы, закрытые специальным защитным покрывальцем –индузием. В спорангиях происходит формирование спор из клетокспорогенной тканив результате редукционного делениямейозом. Спорангии в своем строении имеютножки, в оболочке каждого из них находится ряд специализированных клеток с неравномерно утолщенными стенками –механический поясок(анулюс). При созревании спор клетки механического пояска обезвоживаются первыми, что приводит к их деформации и резкому выворачиванию (раскрытию) спорангия наизнанку с катапультированием спор. Споры, попадая на почву, при благоприятных условиях прорастают впротонему, которая позже разрастается до плоского сердцевидного фотосинтезирующегозаростка– двуполого гаметофита. С нижней стороны гаметофита находятся ризоиды, которыми он крепится к грунту,антеридиииархегонии. Антеридии возникают раньше, ближе к острому краю сердцевидного заростка, а архегонии позже ближе к выемке заростковой пластинки. Разница во времени формирования двух типов гаметангиев способствует перекрестному оплодотворению папоротников. Для перемещения многожгутиковыхсперматозоидовиз антеридиев к яйцеклеткам в архегониях необходима вода, которая скапливается с нижней стороны таллома заростка после дождя или росы. Послеполового процесса(слияния сперматозоида и яйцеклетки) из оплодотворенной яйцеклетки (зиготы) развиваетсязародыш– маленький спорофит, который на первых этапах развития питается за счет гаметофита, а после укоренения переходит на автотрофное питание, становясь независимым организмом. К этому времени гаметофит отмирает.

Рис. 38. Схема жизненного цикла папоротника щитника мужского
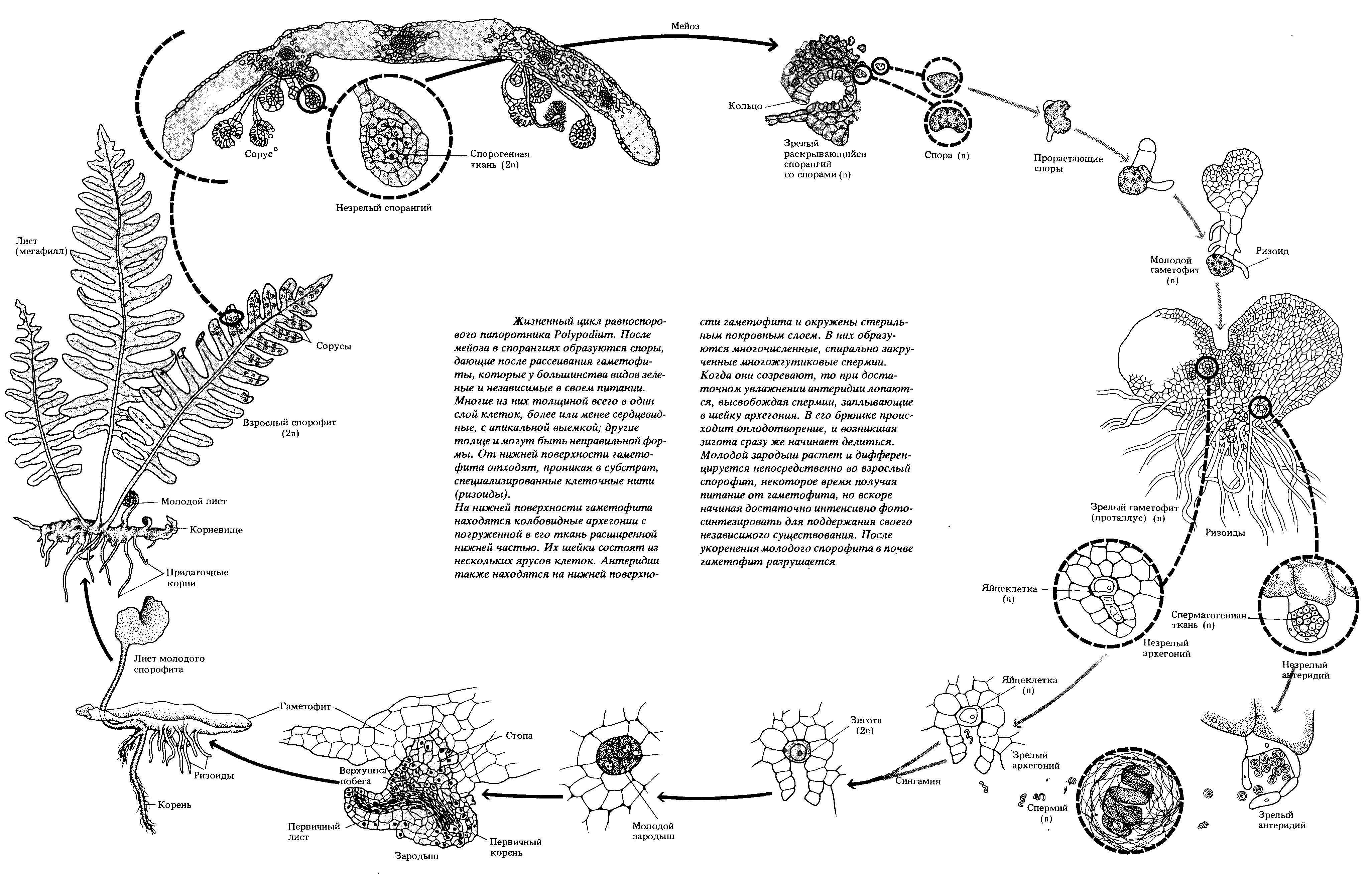
Рис. 39. Жизненный цикл папоротника щитника мужского
Голосеменные растения – группа семенных растений, особенностью которых является наличиесеменных зачатков, расположенных открыто на семенных листках (чешуях). Голосеменные размножаютсясеменамии насчитывают около 1000 видов растений.

